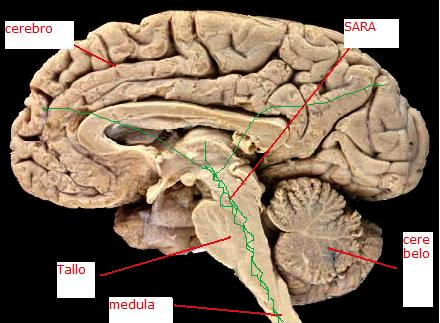
La influencia reticular sobre la corteza puede verse en el encefalograma (EEG) en el hombre o en los animales.
La estimulación de la sustancia reticular ascendente en animales desincroniza el encefalograma, al igual que sucede en el hombre cuando se altera la atención.
Las ondas alfa, son típicas de los estados de relajación.
En el sueño se pueden distinguir 4 estadíos, que se manifiestan en los trazos de las ondas cerebrales. El sueño ligero ligero o superficial se designa como estadío 1 del sueño. El ritmo alfa esta desincronizado y pueden observarse ondas de bajo voltaje de 4 a 6 cps; tambien se hallan presentes esporádicamente ondas delta lentas y de alto voltaje (0 a 3cps).
El estadío 2 del sueño se manifiesta por puntas o descargas de ondas rapidas (14 cps), sobre un fondo de ondas delta con complejos k (una variación negativa aguda, seguida por una onda positiva). En esl estadío 3 del sueño desaparecen las puntas, destaca un ritmo delta y persisten algunos complejos k. En el estadío 4, predominan el el trazado ondas delta de alto voltaje. Los momentos en los que el sujeto sueña pueden detectarse por una combinación del estadío 1 del sueño y movimientos rapidos de los ojos (Rapid eyed movements = REM). Estos últimos son medidos mediante electrodos situados cerca de los músculos oculares.

Sistema Límbico
El sístema límbico junto con el hipotalamo, ha constituido un fecundo campo de estudio del control neural de la emoción. Parte de este sistema han sido incluidas en estudios del aprendizaje.
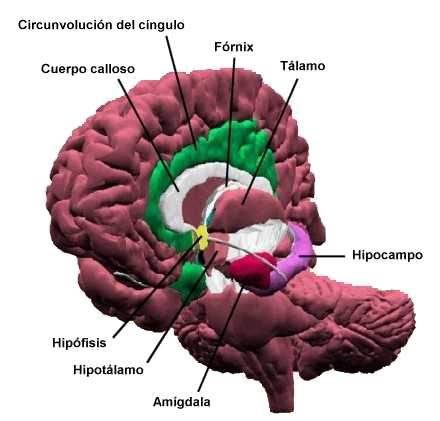
Papez opinaba que los impulsos nerviosos que constituyen la base de las emociones se originan en el hipocampo. a partir de aqui pasarian a los cuerpos mamilares (hipotalámicos) a tráves del fórnix y luego a los núcleos anteriores del tálamo y la circonvolución del cingulo de la corteza cerebral.
Las lesiones del núcleo amigdalino causan docilidad.
Las lesiones septales interfieren con la evitación pasiva.
Las lesiones del hipocampo interfieren con la evitación activa.
El aprendizaje comprende tambien actividad en el sistema limbico. a partir del hipocampo se han registrado ondas kappa como síntoma de aprendizaje con éxito, por ejemplo. Se han aportado datos que muestran que la destrucción del lóbulo temporal (incluyendo al núcleo amigdalino) en el hombre afecta a la capacidad para formar huellas persistentes de la memoria.
El hipocampo interviene en pulsiones, así como en el comportamiento emocional. Los centros ergotropos posteriores controlan el despertar de pulsaciones y las reacciones de furor, mientras que los centros trofotropos anteriores estimulan la saciedad y el reposo.

El metabolismo del Sistema Nervioso Central es elevado y depende de un constante suministro de oxígeno y glucosa.
El sistema vascular del cerebro y de la médula es más extenso que el de cualquier otro órgano del cuerpo. En el hombre, el cerebro recibe cada minuto una cantidad de sangre equivalente a su peso. Si bien, el cerebro representa tan sólo el 2% del peso total del cuerpo, recibe una tercera parte del total de sangre enviado por el corazón a todo el organismo.

El líquido cefalorraquídeo es filtrado a partir de la sangre, en los plexos coroideos de los ventrículos y circula a través de éstos y de los espacios subdural y subaracnoideo.
Este líquido es absorbido por las vellosidades aracnoides y pasa a los senos dúrales.
El líquido cefalorraquídeo ejerce una función amortiguadora, protectora del cerebro.
El liquido intracelular del cerebro se filtra a través de las células gliales que rodean las paredes de los capilares, así como a través de loas propias paredes capilares.
Esta filtración constituye una barrera hematoencefálica; de este modo, el líquido extracelular cerebral no contiene moléculas tan grandes como el líquido extracelular de otros tejidos.

La barreta hematoencefálica evita el paso de la corticosterona, la cual reduce la utilización de glucosa por otras células del organismo, así como del aminoácido esencial que es el ácido glutámico, el cual excita a las células cerebrales.
La barreta hematoencefálica evita el paso de la corticosterona, la cual reduce la utilización de glucosa por otras células del organismo, así como del aminoácido esencial que es el ácido glutámico, el cual excita a las células cerebrales.
Las células nerviosas son adrenérgicos o colinérgicas. En estas últimas, la sustancia transmisora es la acetilcolina y en las primeras, la noradrenalina.
En el hipotálamo las células trofotropas son probablemente colinérgicas y las ergótropas, adrenérgicas.

Información otorgada por: Lizbeth Delgadillo